How does photosynthesis convert light into chemical energy?

The process by which plants, algae, and some bacteria capture the boundless energy streaming from the sun and lock it into the bonds of sugar molecules is one of the most fundamental chemical operations on Earth. This remarkable transformation, known as photosynthesis, is essentially a mechanism for converting light energy into chemical energy. Without this initial conversion, the vast majority of life as we know it would cease to exist, as it forms the base of nearly every food web and is responsible for producing the atmospheric oxygen we breathe. It is a dance involving water, carbon dioxide, and sunlight, orchestrated within the tiny compartments of a plant cell.
# Core Equation
To understand the magic, it helps to first look at the overall recipe. Photosynthesis is an endergonic reaction, meaning it requires an input of energy to proceed. The simplified, overall chemical equation summarizes this energy capture: carbon dioxide and water are transformed, using light energy, into glucose (a sugar) and oxygen.
This equation, while tidy, obscures the intense complexity occurring inside the leaf. The key objective is not just making sugar, but making it using solar power. The initial step must capture that solar energy and convert it into a form that the cell can spend to build those complex sugar molecules—a chemical currency the cell recognizes. The plant doesn't immediately convert sunlight into glucose; rather, it creates short-term, high-energy rechargeable batteries during the first phase of the process.
# Cellular Site
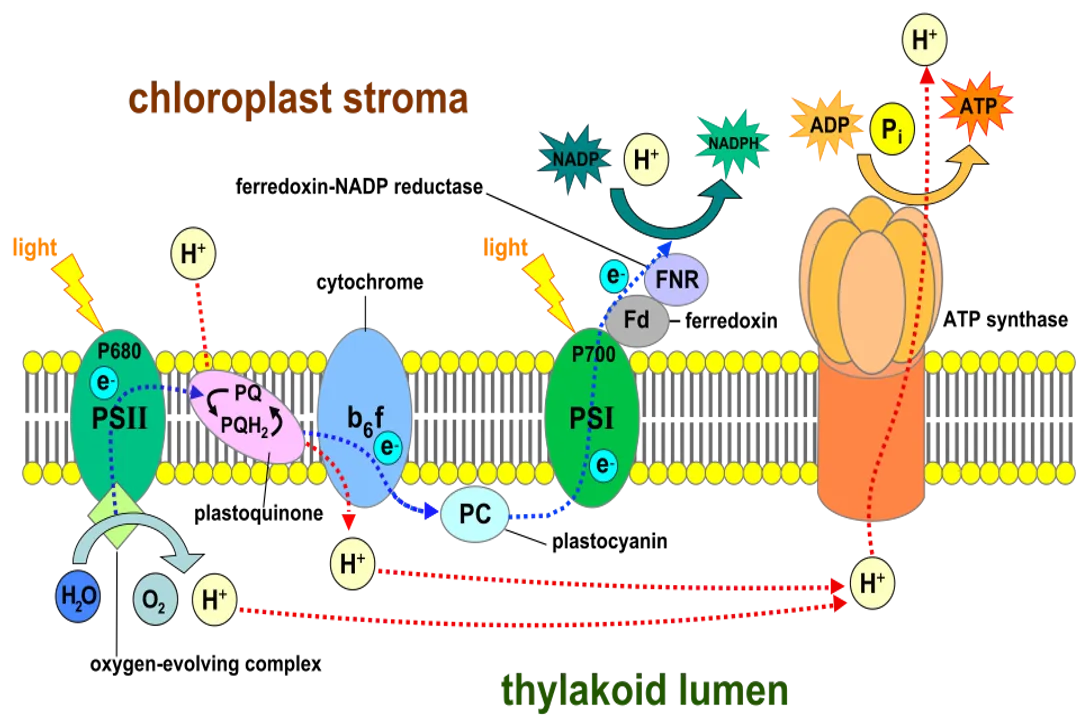
This entire energetic operation takes place primarily within specialized organelles inside plant cells called chloroplasts. Think of the chloroplast as the dedicated solar power factory. Inside this organelle, the structure is highly organized to maximize efficiency. There are two main areas where the two stages of photosynthesis occur: the thylakoids and the stroma.
The thylakoids are flat, interconnected, disc-like sacs or membranes stacked into structures called grana (singular: granum). These membranes contain the pigment that makes plants green, chlorophyll, along with other pigments and proteins organized into functional units called photosystems. This is where the light-capturing machinery resides.
The stroma, conversely, is the fluid-filled space surrounding the thylakoids within the chloroplast. If the thylakoids are the solar panels generating the immediate energy currency, the stroma is the assembly plant where that currency is spent to construct the final product, the sugar. Understanding this spatial separation is crucial, as the products of the light-harvesting stage in the thylakoids must travel to the stroma to fuel the sugar-building stage.

# Light Reactions
The initial phase of photosynthesis, often called the light-dependent reactions, is where the actual conversion of light energy to transient chemical energy occurs. This stage is entirely dependent on the presence of light, as photons—packets of light energy—are the primary input.
# Photon Absorption
The process starts when a photon strikes a pigment molecule, most famously chlorophyll, located within the thylakoid membranes. When chlorophyll absorbs this light, an electron within the pigment molecule gets excited to a higher energy level. This energetic electron is unstable and must immediately pass its energy along. If this energy were simply released as heat or light (fluorescence), the conversion process would fail. Instead, the excited electron is quickly handed off to a neighboring molecule in a highly organized chain of proteins embedded in the thylakoid membrane—this is the electron transport chain (ETC).
# Electron Flow
The movement of this high-energy electron down the ETC is analogous to water flowing downhill, and the cell harnesses that energy of movement. As the electrons move from one protein complex to the next, they release small amounts of energy. This energy is not wasted; it is captured to pump hydrogen ions () from the stroma into the thylakoid lumen (the space inside the thylakoid disc).
This continuous pumping action creates a high concentration of ions inside the lumen compared to the stroma, establishing a powerful electrochemical gradient. This situation is similar to a dam holding back a large volume of water; the potential energy stored in that difference is immense.
# Energy Currency Creation
The cell capitalizes on this stored potential energy through a process called chemiosmosis. The hydrogen ions flow back out of the thylakoid lumen, down their concentration gradient, through a special enzyme complex called ATP synthase. The flow of protons through this enzyme causes it to spin, much like water turning a turbine in a hydroelectric dam. This mechanical action drives the phosphorylation of ADP (adenosine diphosphate) into ATP (adenosine triphosphate). ATP is the universal, short-term energy currency used to power almost all cellular activities.
Simultaneously, the electrons, having given up some of their initial energy, are re-energized by light captured at a second photosystem and are finally used to reduce (nicotinamide adenine dinucleotide phosphate) to its high-energy form, NADPH.
The net result of the light reactions is the conversion of light energy into chemical energy temporarily stored in two molecules: ATP and NADPH. These molecules now bridge the gap between the "light" phase and the "synthesis" phase.
An interesting comparison here is how the inputs are managed. The excited electron must be replaced. This is where water comes in. Water molecules are split in a process called photolysis to provide replacement electrons for Photosystem II. This splitting releases hydrogen ions (contributing to the gradient) and, critically, oxygen () as a waste product. It is a perfect, efficient recycling system where a necessary byproduct () is released, feeding the atmosphere. While most primary sources state the light reactions produce ATP and NADPH, it is worth noting that the Nobel citation mentions that the Nobel Prize in Chemistry in 1988 honored the work elucidating the steps for this photochemical conversion.
# Carbon Fixation
With the immediate energy needs met by ATP and NADPH, the process moves into the second major stage, which occurs in the stroma and is often referred to as the light-independent reactions or the Calvin Cycle. While this stage does not require light directly, it relies entirely on the products (ATP and NADPH) generated during the light reactions, meaning it generally occurs only when the sun is shining.
The purpose of the Calvin Cycle is to take inorganic carbon, in the form of carbon dioxide () from the atmosphere, and convert it into an organic sugar molecule (). This is the step where the final chemical energy is stored in stable, long-term bonds.
# Three Phases
The Calvin Cycle proceeds in three main phases:
Carbon Fixation: A single molecule of enters the cycle and is attached, or fixed, to an existing five-carbon organic molecule called RuBP (ribulose-1,5-bisphosphate). This reaction is catalyzed by the most abundant enzyme on Earth, RuBisCO. The resulting unstable six-carbon molecule immediately splits into two molecules of a three-carbon compound, 3-PGA (3-phosphoglycerate).
Reduction: This is where the energy carriers generated in the thylakoids are finally "spent." ATP supplies the necessary energy, and NADPH supplies the high-energy electrons (reducing power) to convert 3-PGA into a different three-carbon sugar called G3P (glyceraldehyde-3-phosphate). For every six G3P molecules produced in the cycle, one molecule exits the cycle to be used by the plant to build larger carbohydrates like glucose, sucrose, and starch.
Regeneration: The remaining five G3P molecules must be rearranged and recycled to regenerate the initial RuBP acceptor molecules, allowing the cycle to continue capturing more . This regeneration phase also consumes additional ATP energy from the light reactions.
If we consider the entire process, for the plant to net a single molecule of glucose (), the Calvin Cycle must turn six times, fixing six molecules of . This requires a substantial investment of the captured light energy: 18 ATP molecules and 12 NADPH molecules are consumed for the synthesis of one glucose unit. This illustrates how light energy, captured first in ATP/NADPH, is slowly and methodically stitched into the stable energy bonds of the sugar molecule.

# Energy Transfer Comparisons
It is essential to distinguish between the type of energy being converted at each stage. The initial step is a photochemical conversion where electromagnetic energy (light) is transformed into kinetic energy (excited electrons) and then into the potential energy carried by the chemical bonds of ATP and NADPH. These are high-energy, short-lived carriers. The second stage, the Calvin Cycle, is a thermodynamic conversion, using the energy stored in ATP and NADPH to build high-energy, long-lasting carbon bonds, yielding complex carbohydrates.
One interesting comparison arises when looking at efficiency and structure. Plants have optimized this system over eons. For instance, the efficiency with which the light-harvesting complex captures photons is nearly perfect, ensuring very little initial energy is lost unnecessarily. However, the enzyme RuBisCO, used in the carbon fixation step, is notoriously slow and prone to making errors (fixing oxygen instead of , a process called photorespiration). This suggests an evolutionary trade-off: the initial light capture mechanism is incredibly precise, but the carbon-building step is constrained by an older, less perfect catalyst, which is why some specialized plants (like C4 or CAM plants) have evolved supplementary pathways to concentrate and bypass RuBisCO's inefficiency in hot, dry conditions.
To illustrate the resource cost, consider a hypothetical scenario: if a patch of common temperate grass fixes enough carbon in a day to produce of biomass, it has consumed approximately of water and the energetic equivalent of about $4.7$ liters of gasoline (in terms of potential energy converted) during that time, highlighting the immense energetic investment required to move from atmospheric gas to solid structure.
# Global Energy Flow
Photosynthesis is not merely a process occurring within a single leaf; it dictates the energy dynamics of the entire biosphere. Life on Earth is fundamentally powered by this conversion of radiant energy. Producers, those organisms capable of photosynthesis, capture light and transform it, making that energy available to consumers (herbivores) and then secondary consumers (carnivores).
Furthermore, the output of the light reactions—oxygen—is the byproduct that made complex aerobic life possible. Before photosynthetic organisms evolved, Earth's atmosphere had very little free oxygen. The evolution of oxygenic photosynthesis fundamentally changed the planet's chemistry and allowed for the evolution of respiration, the process that releases the chemical energy stored by photosynthesis when needed.
The significance of this conversion lies in its dual role: energy storage and matter cycling. It stores energy by creating C-H bonds in sugars, and it cycles matter by pulling carbon from the atmosphere and integrating it into the living world. This continuous flow and conversion, powered by the sun, maintains the delicate chemical balance necessary for life. The mechanism, while complex in its intermediate steps involving electron transport and carbon fixation, adheres strictly to the laws of thermodynamics, managing energy transfer from one form to another with high, though imperfect, fidelity.
# Pigments and Light Capture
The efficiency of the initial light conversion hinges on the light-absorbing molecules, the pigments. Chlorophyll is the primary pigment, directly driving the light reactions, while chlorophyll and carotenoids act as accessory pigments. These accessory pigments are essential because they absorb wavelengths of light that chlorophyll misses and then pass that captured energy over to chlorophyll within the reaction centers of the photosystems. This collection strategy broadens the spectrum of usable sunlight, effectively increasing the overall capture rate of the solar energy available to the plant.
# Conclusion on Conversion
The journey from a fleeting photon to a stable sugar molecule is a two-act play unfolding within the chloroplast. The first act, in the thylakoids, is a rapid, high-energy physical process where light energy is immediately transduced into electrical energy (moving electrons) and then into the readily usable chemical potential of ATP and NADPH. The second act, in the stroma, is a slower, cyclical construction project where that temporary chemical potential is painstakingly invested to reduce atmospheric carbon dioxide into the dense, stable chemical bonds of glucose. Photosynthesis is the master conversion technology of nature, packaging the energy of a star into the structure of a leaf.
Related Questions
#Citations
Photosynthesis - National Geographic Education
Photosynthesis - The Cell - NCBI Bookshelf - NIH
Harnessing Sunlight: How Plants Convert Light to Chemical Energy
Photosynthesis: Chemical energy from light - Nobel Prize
Photosynthesis Converts Solar Energy Into Chemical ... - Ask Nature
Photosynthesis - AGU
Intro to photosynthesis (article) - Khan Academy
A Primer on Photosynthesis and the Functioning of Cells
3.4: Energy Enters Ecosystems Through Photosynthesis
Photosynthesis - Wikipedia