How do stem cells maintain pluripotency?
The remarkable ability of pluripotent stem cells (PSCs) to indefinitely self-renew while holding the potential to become any cell type in the body is governed by an intricate balance of internal genetic programs and external environmental signals. This state of plasticity, crucial for regenerative medicine, is far from static; it is actively maintained through precise molecular management, distinguishing between the established "primed" state found in typical human lines and the more immature "naive" state.
# Core Network
At the heart of maintaining this undifferentiated state is a core regulatory circuit composed primarily of the transcription factors Octamer-binding transcription factor 4 (OCT4), SRY (sex-determining region Y)-Box 2 (SOX2), and NANOG. These factors engage in auto-regulatory and cross-regulatory loops, binding to hundreds of promoters to actively transcribe genes necessary for pluripotency while simultaneously repressing gene expression associated with differentiation.
The requirement for each component is highly specific. OCT4 and SOX2 often cooperate to regulate the general pluripotent program. NANOG acts to stabilize this state, with its loss often leading to spontaneous differentiation toward specific lineages, like primitive endoderm in some contexts. Historically, c-Myc was considered essential in the initial reprogramming cocktail (OSKM), but research shows it is dispensable for the maintenance of pluripotency itself, primarily acting to enhance the efficiency and speed of the transition from a somatic cell by promoting chromatin accessibility.
It is interesting to observe that the sheer number of transcription factors binding a promoter often correlates with gene activity in the pluripotent state; promoters bound by four or more core factors are much more likely to be actively transcribed. This combinatorial binding ensures that the entire regulatory landscape remains tilted away from any single lineage commitment.
# External Cues
While the core factors dictate what pluripotency is, external growth factors dictate how it is kept stable in vitro—a necessity because pluripotency is inherently unstable and primed for differentiation. The required signals, however, differ significantly between species, which points to a convergent functional outcome achieved through divergent pathway reliance.
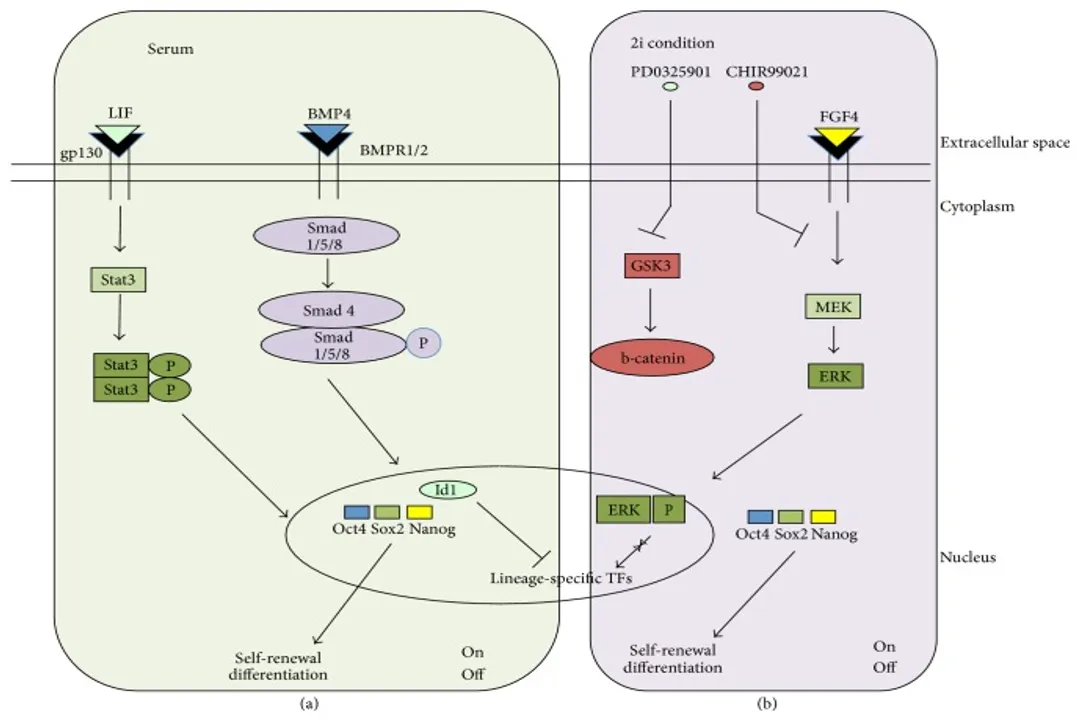
Mouse Embryonic Stem Cells (mESCs) are traditionally maintained by Leukaemia Inhibitory Factor (LIF), which signals through JAK/STAT3, often synergizing with Bone Morphogenetic Proteins (BMPs) that suppress the differentiation-driving MAPK/ERK pathway. In contrast, human Embryonic Stem Cells (hESCs) do not rely on LIF for self-renewal; instead, they require Fibroblast Growth Factor 2 (FGF2), Activin A/Nodal, and sometimes Insulin-like Growth Factor (IGF) signaling. Activin/Nodal activates SMAD2/3 signaling, which cooperates with SMAD2/3 target binding at pluripotency genes like NANOG.
A deeper look reveals this disparity: mESCs in standard serum/LIF conditions are metastable, sitting between the true "ground state" and epiblast-like cells. When mESCs are pushed into the naive ground state using inhibitors (2i) that block MEK/ERK and GSK3 signaling, they become independent of the LIF/STAT3 pathway. This suggests that the purpose of the extrinsic signals is to block specific differentiation pathways (like ERK activation in mouse or BMP activation in human) that would otherwise push the cell out of the pluripotent window. It seems the universal mechanism is not a specific activated pathway, but rather the active suppression of competing differentiation cues that are inherent to the cell’s environment or history.

# Chromatin Landscape
Maintenance of pluripotency requires that differentiation genes are silenced, but in a manner that allows for rapid activation upon receiving the right cue. This is managed through epigenetic control, where ES cell chromatin is generally euchromatic.
A key feature is the presence of bivalent domains—genomic regions marked simultaneously with the active mark H3K4me3 and the repressive mark H3K27me3. These domains often reside over developmental transcription factors that are poised for activation, meaning the necessary machinery is ready to deploy quickly upon commitment. The repressive H3K27me3 mark is largely set by the Polycomb Repressive Complex 2 (PRC2), which helps keep alternative fates quiet without permanently shutting them down via irreversible heterochromatin.
Furthermore, the chromatin structure in PSCs is described as hyperdynamic or "breathing," characterized by rapid turnover of histone proteins like H2B and H3 compared to differentiated cells. Preventing this dynamic exchange by stabilizing the structure can actually block differentiation and lead to cell death in prolonged culture.
In contrast to histone modifications, DNA methylation at CpG islands appears less critical for maintaining the naive state itself, as DNA methylase-deficient cells can self-renew. However, DNA methylation does play a role in repressing differentiation-associated genes in the overall genome. Conversely, when cells are reprogrammed from a somatic state, the erasure of repressive marks like H3K9me3 is a major barrier that must be overcome for the core pluripotency genes to become accessible to the OSKM factors. This highlights a crucial functional difference: maintenance involves fine-tuning the bivalent state and dynamic turnover, while induction requires a massive dismantling of somatic-memory heterochromatin.
# Metabolic States
The energetic state of the cell is also rewired during the acquisition and maintenance of pluripotency. Somatic cells rely primarily on oxidative phosphorylation (OXPHOS) in their mitochondria to generate ATP. However, PSCs, mimicking pre-implantation embryos, undergo a metabolic shift to prioritize anaerobic glycolysis (the Warburg effect).
This shift results in mitochondria with less-defined cristae and lower net ATP production, instead relying on lactate production. This metabolic change is linked to activation of the hypoxia-inducible pathway to compensate for the reduced mitochondrial efficiency. The metabolic state is so closely tied to identity that manipulating the availability of certain nutrients can affect the epigenetic state; for instance, threonine depletion reduces S-adenosyl-methionine (SAM) activity, leading to reduced H3K4 trimethylation and subsequent differentiation. L-Proline, another amino acid, can act as a signaling molecule that remodels methylation profiles (H3K9 and H3K36), potentially inducing a transitional state between naive mESCs and epiblast stem cells (EpiSCs).

# States and Plasticity
The maintenance mechanisms also explain the critical difference between naive and primed pluripotency, a distinction essential for understanding human versus mouse cell biology.
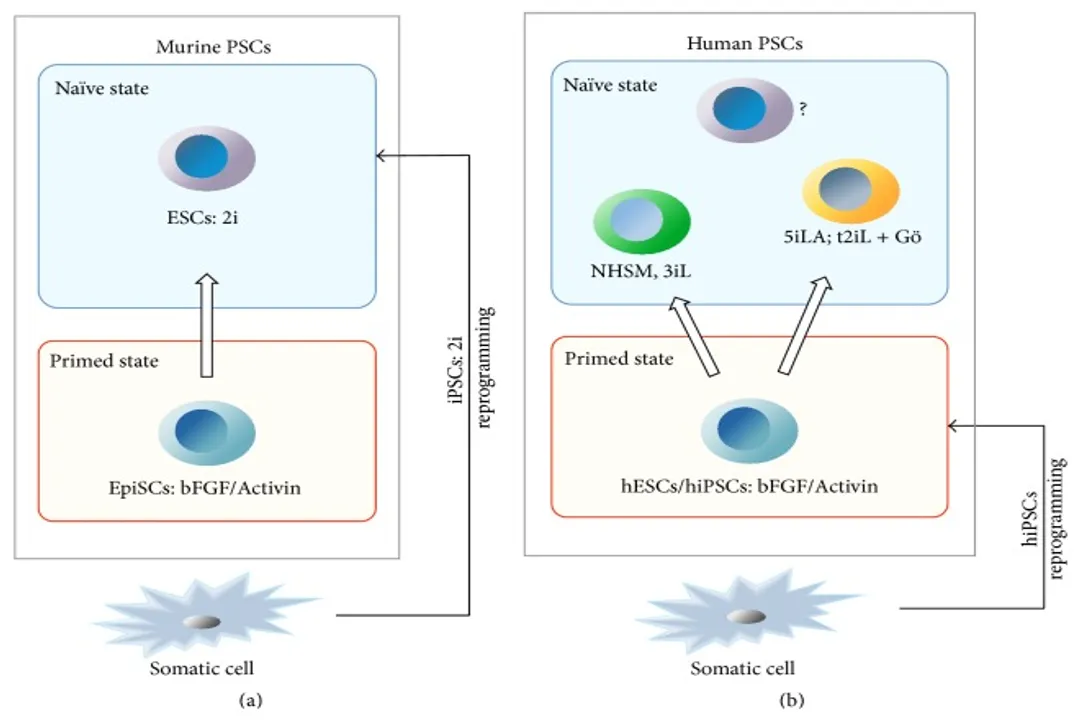
Naive Pluripotency: This state is considered the most immature, resembling the Inner Cell Mass (ICM) of the pre-implantation embryo. Murine ESCs cultured in 2i/LIF (which inhibits MEK/ERK and GSK3) achieve a robust "ground state" naive pluripotency, which is highly homogenous and can be derived from any mouse strain. Naive cells exhibit higher chimera contribution ability and hypomethylation.
Primed Pluripotency: This state is more developmentally advanced, resembling the post-implantation epiblast (EpiSCs in mice). Conventional hESCs and hiPSCs, maintained using factors like FGF2 and Activin A, are generally considered primed. Primed cells grow as flat colonies, are often dependent on JAK/STAT3-independent pathways (like SMAD2/3 for NANOG), and importantly, female cells already exhibit X-chromosome inactivation (XCI), an epigenetic hallmark of commitment, unlike naive mESCs which maintain two active X chromosomes (XaXa).
The existence of the human naive state, which more closely mirrors the mouse ground state, has been the subject of intense refinement, requiring specific chemical cocktails to induce or maintain it. The fact that different optimized media (like NHSM, 3iL, 5i/L/A, or t2iL + Gö) yield slightly different transcriptional profiles suggests that the true human ground state remains elusive, or that stability in culture is highly dependent on the exact combination of signaling suppressors used to mimic the in vivo early embryonic niche.
The entire process of maintaining pluripotency is therefore a careful, active work of suppression: suppressing differentiation pathway activation via specific extrinsic signals, suppressing lineage-specific gene expression via poised bivalent chromatin, and maintaining a glycolytic metabolism to support rapid cell cycling without commitment. The cell must continually exert control to resist the powerful epigenetic memory pushing it toward its differentiated somatic origin. A fascinating consequence of this delicate balance is observed in the phenomenon where even partially reprogrammed cells (iRSCs) may resume the process depending on cell density, indicating that cell-cell communication—a form of extrinsic signaling—can act as a critical checkpoint for progression toward the stabilized pluripotent state.
Related Questions
#Citations
Pluripotent Stem Cells: Current Understanding and Future Directions
Understanding pluripotency—how embryonic stem cells keep their ...
Pluripotent stem cells: Biology and applications - Abcam
Pluripotency maintenance mechanism of embryonic stem cells and ...
Pluripotent stem cells: induction and self-renewal - Journals
A method to maintain and propagate pluripotent human ES cells