How do membranes regulate cellular transport?
The cell membrane, a thin boundary surrounding every living cell, is far more than just a static fence; it is a dynamic, selective gatekeeper that dictates precisely what enters and leaves the internal cellular environment. [5][7] This regulation is fundamental to life, ensuring that essential nutrients are imported, waste products are expelled, and the delicate internal balance, known as homeostasis, is maintained. [4][9] The capacity for this precise control rests almost entirely on the structure and composition of the membrane itself and the specialized components embedded within it. [3]
# Bilayer Makeup
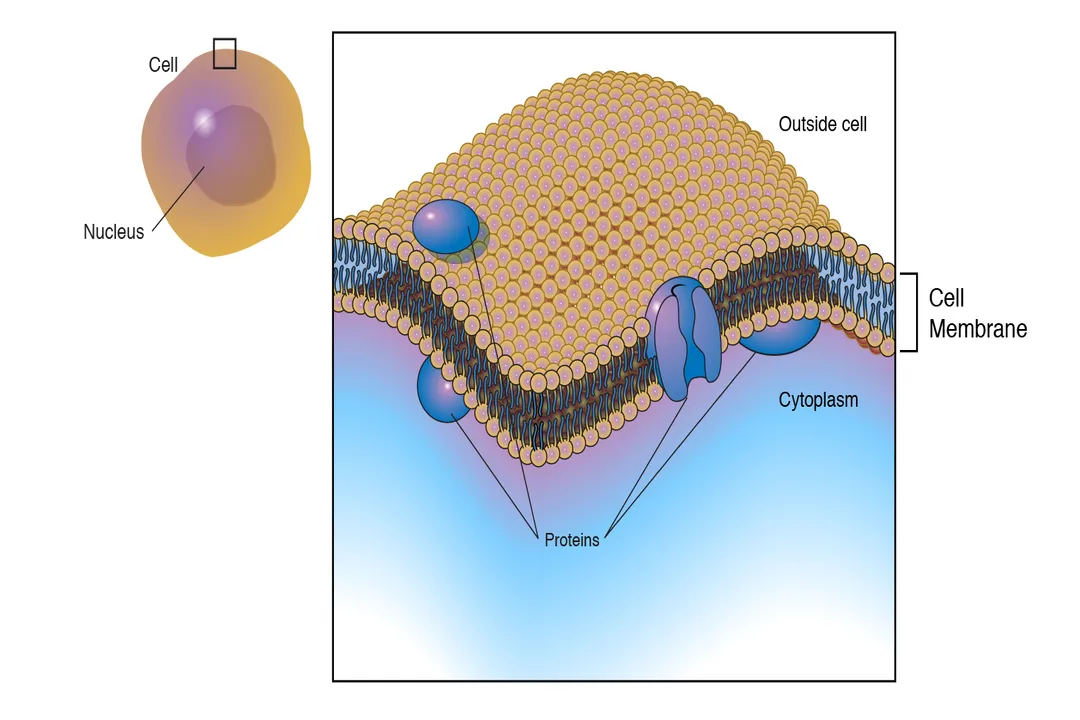
The foundational structure permitting this regulation is the phospholipid bilayer. [5] Phospholipids are amphipathic molecules, meaning they possess both hydrophilic (water-loving) heads and hydrophobic (water-fearing) tails. [5] In an aqueous environment, like the body's interior, these molecules spontaneously arrange themselves into two layers, with the heads facing outward toward the water on both sides and the tails tucked safely in the middle. [5] This arrangement creates a barrier that is inherently impermeable to most charged molecules, large polar substances, and ions, as they cannot easily pass through the fatty, nonpolar core. [6][9]
This fundamental barrier gives rise to the first major category of transport: passive transport. Passive processes require no direct input of cellular energy, typically Adenosine Triphosphate (ATP), because they rely on the inherent tendency of molecules to move down their concentration gradient—from an area of higher concentration to an area of lower concentration. [6][9]
# Passive Movement
Several types of passive transport exist, each suited to different types of materials trying to cross the membrane. [1]
# Simple Diffusion
The simplest mechanism is simple diffusion, which applies to very small, nonpolar molecules like oxygen () or carbon dioxide (), and some small uncharged polar molecules like water in some contexts. [1][6] These molecules move directly through the lipid bilayer in response to their concentration gradient. [6] Imagine a crowded room where a door is opened slightly; people will naturally flow out toward the less crowded area until the density is equalized. Similarly, the movement of a solute across the membrane continues until equilibrium is reached, at which point the net movement stops, though molecules continue to move randomly back and forth. [6]
# Osmosis Water Movement
Water movement warrants special attention because of its sheer volume and critical role in cell volume regulation; this process is called osmosis. [1][9] Osmosis is specifically the diffusion of water across a selectively permeable membrane. [1] If a cell is placed in a solution with a higher solute concentration outside than inside (a hypertonic solution), water will move out of the cell to try and dilute the external environment, causing the cell to shrink. [1] Conversely, in a hypotonic solution (lower external solute concentration), water rushes into the cell, potentially causing it to swell and even burst, a process called lysis. [1]
# Mediated Transport
While small nonpolar molecules slip right through, many essential substances—like ions, sugars, and amino acids—are too large or too polar to pass the lipid core effectively, even if moving down their gradient. [6][9] These materials require assistance from transport proteins, a process known as facilitated diffusion. [1][6]
Facilitated diffusion uses specific membrane proteins to move substances down their concentration gradient, much like simple diffusion, but faster. [6]
- Channel Proteins: These proteins form hydrophilic pores or tunnels through the membrane. [1] Many are gated, meaning they open or close in response to specific signals, such as changes in voltage (voltage-gated channels) or the binding of a specific chemical messenger (ligand-gated channels). [6] They allow very rapid transport of specific ions like sodium () or potassium (). [1][6]
- Carrier Proteins: These proteins bind to the specific molecule they transport—like a key fitting a lock—undergo a conformational (shape) change, and then release the molecule on the other side of the membrane. [1][6]
A key aspect of passive transport is its saturation kinetics. Because facilitated diffusion relies on a finite number of available carrier or channel proteins, if the concentration of the substance needing transport becomes extremely high, all the protein carriers become occupied. At this point, increasing the external concentration further does not increase the transport rate, as the rate is limited by how quickly the proteins can flip back and forth; they become saturated. [3]
Consideration for Cellular Energetics: When we compare simple diffusion to protein-mediated passive transport, it’s interesting to note the trade-off between speed and reliance on structure. Simple diffusion is slow but requires nothing more than the intact lipid bilayer. Facilitated diffusion is immensely faster—sometimes millions of times faster for ions—but requires the synthesis, proper insertion, and maintenance of complex protein machinery. A cell heavily reliant on rapid metabolite exchange must dedicate significant resources to synthesizing these proteins, a resource cost not present when simply waiting for oxygen to diffuse across the membrane.

# Active Transport Needs
While passive transport handles the simple influx and efflux driven by concentration differences, sometimes the cell needs to move something against its concentration gradient—from a low concentration area to a high concentration area. This is akin to pumping water uphill; it requires an energy investment, thus classifying it as active transport. [6][9]
# Primary Active Transport
Primary active transport directly couples the movement of a substance against its gradient to an energy-releasing process, most commonly the hydrolysis of ATP. [6][9] The most famous example is the sodium-potassium pump ( ATPase). [6][9] This pump is critical for maintaining the electrical potential across the membrane and cell volume. [6] For every cycle, it uses one ATP molecule to actively pump three sodium ions out of the cell and two potassium ions into the cell. [6][9] This creates both a concentration gradient (more outside, more inside) and an electrical gradient (the outside becomes more positive relative to the inside). [6]
Other primary active transporters include proton () pumps, which are vital in organelles like the lysosomes to maintain acidic internal pH, and calcium () pumps. [9]
# Secondary Active Transport
Secondary active transport, also known as co-transport, gets its energy indirectly from an existing electrochemical gradient generated by primary active transport. [6][9] Instead of using ATP directly, the movement of a substance down its steep electrochemical gradient (often moving back into the cell, following the pump's work) is coupled with the movement of another substance up its own gradient. [6][9]
There are two types of co-transport:
- Symport: Both molecules move in the same direction across the membrane. [6] For example, the /Glucose symporter uses the inward rush of to drag glucose into the cell, even when glucose concentration inside is already high. [6]
- Antiport: The molecules move in opposite directions. [6] The antiporter is a classic example, using the energy of entering the cell to pump out against its gradient. [6]
# Bulk Movement Processes
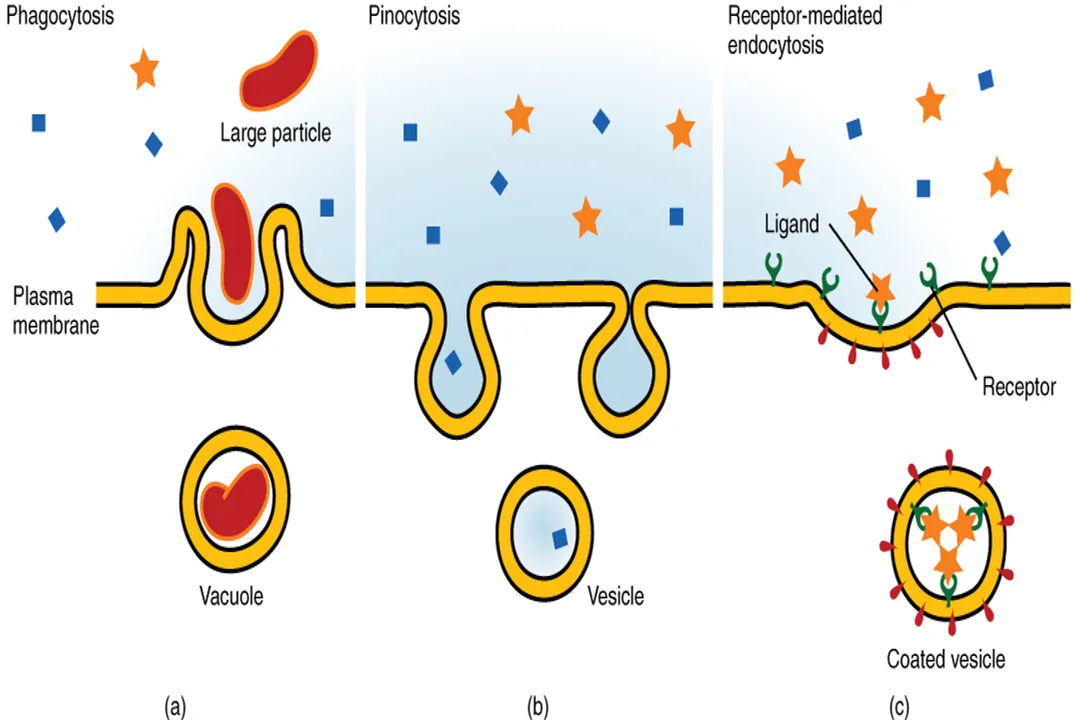
When cells need to move very large particles, macromolecules, or significant quantities of substance that cannot pass through individual protein channels or carriers, they employ mechanisms that involve physically reshaping the plasma membrane itself: bulk transport. [1] These processes are always forms of active transport, requiring significant energy and involving vesicle formation. [1]
# Moving Things In
The process of bringing material into the cell via the membrane engulfing it into a vesicle is called endocytosis. [1]
- Phagocytosis ("Cell Eating"): The cell extends its pseudopods to surround and engulf a large particle, such as bacteria or cellular debris, forming a large vesicle called a phagosome. [1] This is crucial for immune cells like macrophages.
- Pinocytosis ("Cell Drinking"): The cell takes in small droplets of extracellular fluid and any dissolved solutes by forming smaller vesicles. [1] This is a non-specific sampling mechanism.
- Receptor-Mediated Endocytosis: This is highly specific. Receptors on the cell surface bind only to specific target molecules (ligands). Once a sufficient number of receptors are clustered, the area invaginates to form a vesicle containing only those specific molecules. [1] This is how cells can quickly import large amounts of cholesterol, for instance. [1]
# Moving Things Out
Conversely, exocytosis is the process where intracellular vesicles fuse with the plasma membrane, releasing their contents into the extracellular space. [1] This is the primary pathway for secreting hormones, neurotransmitters, and waste products that are too large to be pumped out individually. [1]
# Protein Specialization
The sheer variety of transport methods highlights that the function of the membrane is dictated by its protein components, which account for much of its functional complexity. [3] The phospholipid bilayer provides the basic structure, but the embedded proteins provide the gatekeeping and pumping capabilities. [3]
The different types of proteins involved can be broadly categorized based on their function:
| Protein Type | Primary Function | Energy Requirement | Mechanism Example |
|---|---|---|---|
| Pumps (ATPases) | Establish gradients by moving substances against their gradient [6] | Direct ATP (Primary Active) | Pump [9] |
| Channels | Form selective pores for rapid, passive ion movement [1][6] | None (Passive) | Voltage-gated channel [6] |
| Carriers/Transporters | Bind a solute, change shape, and move it across (can be passive or active) [6] | Passive or Coupled to a Gradient (Secondary Active) | Glucose transporter [6] |
Editor's Analysis: It is worthwhile to consider the concept of transport stoichiometry when analyzing cellular efficiency. Primary active transporters like the pump operate with a specific ratio (). This imbalance is not accidental; it is engineered to make the cell interior electrically negative relative to the outside. This established potential energy difference is then harvested by secondary active transporters to drive nutrient uptake. If a cell were to suddenly lose its ability to pump out effectively, not only would the gradient collapse, but the driving force for nearly all secondary active nutrient import would vanish instantly, demonstrating a cascading failure of regulatory systems.
# Membrane Fluidity and Transport Rate
While we focus on the proteins doing the work, the physical state of the lipid bilayer itself plays a supporting, yet vital, role in transport regulation. [5] The membrane is not rigid; it is described as a fluid mosaic, meaning the lipids and proteins are generally able to move laterally within the plane of the membrane. [5]
This fluidity directly impacts the efficiency of protein-mediated transport. For a carrier protein to complete a cycle—binding a molecule on one side, flipping its conformation, and releasing it on the other—it must be able to physically move and change shape within the lipid sea. [3] If the surrounding lipids become too stiff (e.g., due to high cholesterol content or low temperature), the conformational changes required by carrier proteins slow down drastically. This effectively decreases the maximum transport rate () for those specific carriers, even if the concentration of the substrate remains high. Therefore, regulating membrane fluidity is an indirect, yet essential, way for a cell to control its rate of nutrient acquisition and waste disposal, acting as a fine-tuning dial for the entire transport machinery. [5]
In summary, cellular transport regulation is a multi-layered system. It begins with the passive permeability barrier of the lipid bilayer, is greatly enhanced by specific protein channels for rapid, gradient-driven movement, and is ultimately controlled by energy-dependent pumps that build the necessary electrochemical gradients. The fine coordination between these passive and active systems ensures the cell maintains its unique internal composition, distinguishing it fundamentally from its surroundings. [4][7]
#Videos
In Da Club - Membranes & Transport: Crash Course Biology #5
Related Questions
#Citations
Transport across cell membrane - BYJU'S
In Da Club - Membranes & Transport: Crash Course Biology #5
Membrane Transport - Chemistry LibreTexts
Confused about the transport of materials across cell membranes
Cell Membrane (Plasma Membrane)
Transport across a cell membrane - Khan Academy
Membrane transport - Wikipedia
Transport Across Cell Membranes - Free Sketchy MCAT Lesson
Cell Transport – Interactive Anatomy and Physiology